

甾醇是广泛存在于生物体内的一类天然活性成分,具有抗肿瘤、调节激素水平、促进新陈代谢和预防心血管系统疾病等功效。甾醇是生物膜的重要组成成分之一,通过将其侧链嵌入膜的疏水中心限制脂肪酰基链的活动,从而有效调节膜的流动性和渗透性[1]。植物甾醇种类繁多、功能多样,在植物的生长发育过程中发挥重要作用[2-3]。目前自然界中已发现了近300种植物甾醇,其中菜油甾醇、β-谷甾醇和豆甾醇是大多数植物中主要的甾醇[3]。植物甾醇的主要特点是含有环戊烷多氢菲母核以及C17位连有不同的烷烃侧链,多数植物甾醇的C24位具有甲基或乙基。根据C24位烷基类型不同,可将植物甾醇分为C24甲基甾醇和C24乙基甾醇,前者以菜油甾醇为典型代表,后者以β-谷甾醇和豆甾醇为典型代表[3-4]。
植物甾醇在植物体内以游离甾醇和共轭甾醇的形式存在,共轭甾醇根据C3位羟基结合的不同基团又分为甾醇酯、甾醇糖苷和酰基甾醇糖苷[3]。甾醇酯在膜甾醇稳态中发挥核心作用,可以将细胞膜的游离甾醇维持在正常生理水平。同时,甾醇酯兼具植物甾醇与脂肪酸的双重功效,与游离甾醇相比具有更好的脂溶性和降胆固醇的效果[5-6]。甾醇糖苷和酰基甾醇糖苷在质膜的脂筏中高度富集,在跨膜信号传递中起重要作用[7]。
大多数植物中甾醇糖苷和酰基甾醇糖苷的含量相对较低,通常少于总甾醇的20%,但番茄果实中的甾醇糖苷和酰基甾醇糖苷占总甾醇的85%以上[8-10]。共轭甾醇可参与植物因冷热和干旱等非生物胁迫引起的应激反应[7],冷胁迫和干旱胁迫导致拟南芥Arabidopsis thaliana (L.) Heynh中甾醇糖苷和酰基甾醇糖苷的浓度增加[11],苹果树皮中酰基甾醇糖苷的浓度增加[12]。油菜素内酯是菜油甾醇的衍生物,也是继生长素、赤霉素、细胞分裂素、脱落酸和乙烯之后发现的第6大类植物激素,可调节细胞伸长、细胞分裂、光形态发生、木质部分化和繁殖等多种过程,参与非生物和生物胁迫反应,对植物的生长发育具有重要的调控作用[13]。
植物甾醇具有降胆固醇、降血糖和抗炎等多种药理活性,是一类对人体非常有利的次生代谢物质,被誉为“生命的钥匙”[14]。植物甾醇可通过2个途径来降低体内胆固醇浓度:一方面,因其结构与胆固醇相似,在肠道内会竞争和阻碍胆固醇的吸收[15-16];另一方面,植物甾醇可以与肠道中的胆固醇共结晶并通过粪便排出体外[17]。因此,摄取植物甾醇可降低血液中总胆固醇的水平以及低密度脂蛋白胆固醇的浓度,降低心血管疾病患病风险。妊娠期糖尿病已成为全世界孕妇的严重健康风险之一,每日食用富含植物甾醇的人造黄油可改善妊娠期糖尿病患者的胰岛素抵抗和高血脂的症状[18]。在小鼠饮食中加入适量的β-谷甾醇和豆甾醇可以有效改善由于葡聚糖硫酸钠诱导引起的结肠炎[19],且豆甾醇比β-谷甾醇表现出更好的抗急性结肠炎活性。
人体自身无法合成植物甾醇,体内的植物甾醇都是通过食用植物油、水果、坚果、谷物和豆类等食物摄取的。当前植物甾醇主要从植物中提取,但提取过程中存在甾醇含量低、提取步骤繁琐、污染环境等弊端。近年来,在微生物中构建植物天然产物的生物合成途径,并通过发酵生产植物天然产物具有生产周期短、发酵产物专一、减少对农业土地的竞争、易于分离纯化等优势,是一种高效绿色的生产模式。构建植物甾醇人工生物合成体系的前提是要解析其生物合成途径,多年来在这方面取得了许多研究进展。本文对植物甾醇及其主要衍生物的生物合成途径进行归纳总结,以期为利用合成生物学技术构建生产植物甾醇的微生物细胞工厂提供理论支持。
1 植物甾醇的生物合成途径
植物甾醇的生物合成途径可分为3个阶段:乙酰辅酶A到环阿屯醇的合成、环阿屯醇到4-甲基-24-亚甲基胆甾-7-烯醇的合成和24-亚甲基胆甾-7-烯醇到菜油甾醇、β-谷甾醇和豆甾醇的合成(图1)。前2个阶段是所有植物甾醇合成的共有部分,植物中以环阿屯醇为节点,一部分代谢流通往C24-烷基植物甾醇通路,另一部分代谢流则走向胆固醇合成通路。第3个阶段以24-亚甲基胆甾-7-烯醇为节点,分为C24-甲基甾醇合成途径(合成菜油甾醇)和C24-乙基甾醇合成途径(合成β-谷甾醇和豆甾醇)。
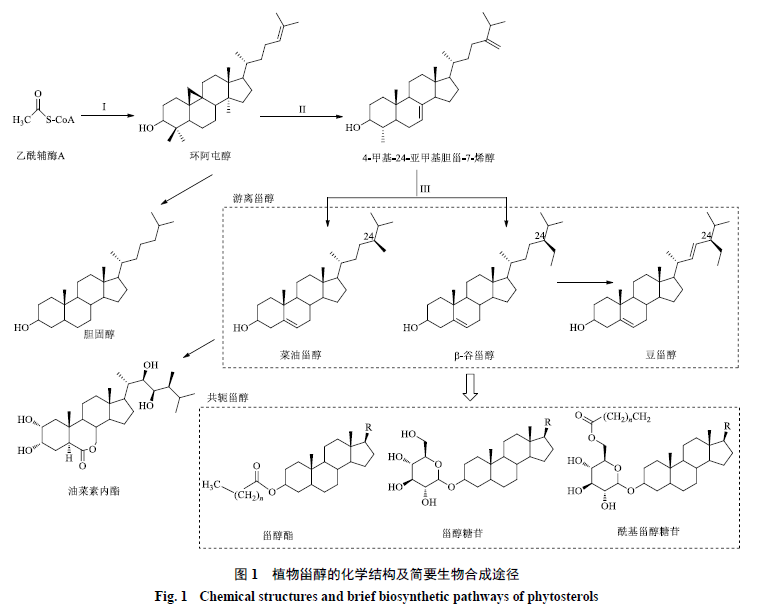
1.1 乙酰辅酶A到环阿屯醇的生物合成途径
在环阿屯醇的生物合成中,经甲羟戊酸途径合成的异戊烯二磷酸(isopentenyl diphosphate,IPP)和二甲基烯丙基二磷酸(dimethylallyl diphosphate,DMAPP)首先经过香叶基二磷酸合酶(geranyl diphosphate synthase,GPPS)和法尼基二磷酸合酶(farnesyl diphosphate synthase,FPPS)的催化,转化为香叶基二磷酸(geranyl diphosphate,GPP)和法呢基二磷酸(farnesyl diphosphate,FPP)。之后,2个FPP分子在角鲨烯合酶(squalene synthase,SQS)的作用下以头-头相接的方式缩合生成角鲨烯,后者在角鲨烯环氧化酶(squalene epoxidase,SQE)的作用下被氧化为2,3-环氧角鲨烯。最后,2,3-环氧角鲨烯经过环阿屯醇合成酶(cycloartenol synthase,CAS)的催化作用生成环阿屯醇(图2)。
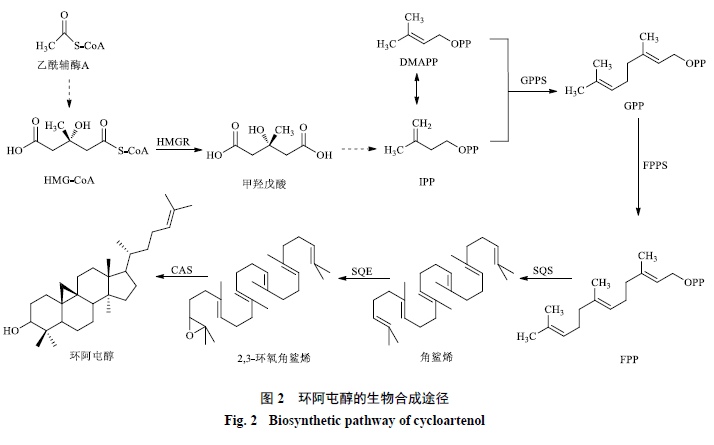
3-羟基-3-甲基戊二酰辅酶A还原酶(
3-hydroxy-3-methylglutaryl-CoA reductase,HMGR)催化三羟基三甲基戊二酸单酰辅酶A转化为甲羟戊酸,是甲羟戊酸途径的限速酶[20-21]。近年来,越来越多的证据表明HMGR在植物甾醇产量提升方面起着重要作用。川贝母Fritillaria cirrhosa D. Don中HMGR(FcHMGR)完整蛋白不能在大肠杆菌中表达,而N端截短后的FcHMGR可以在大肠杆菌中成功表达,体外实验也证明截短的FcHMGR具有还原酶活性[22]。将人参Panax ginseng C. A. Meyer中的PgHMGR1基因在拟南芥中过表达,突变植株中β-谷甾醇含量提高了2倍,菜油甾醇和环阿屯醇的含量均提高了1.8倍[23],证实了人参中HMGR基因在植物甾醇产量提升方面起重要调节作用。
SQS催化2个FPP分子缩合生成角鲨烯的反应分2步进行。第1步,2个FPP分子头对头偶联形成1个稳定的环丙基羰基二磷酸中间体,即前角鲨烯二磷酸(presqualene diphosphate,PSPP);第2步,PSPP的环丙烷环在还原型烟酰胺腺嘌呤二核苷酸磷酸(nicotinamide adenine dinucleotide phosphate,NADPH)的还原作用下被打开,碳骨架发生重排生成角鲨烯[24]。
据报道,去除疏水性C末端部分可以促进可溶性重组蛋白的表达[25],Dai等[26]在越南油茶Camellia vietnamensis Huang中SQS的C端删除了第386~408个氨基酸,将截短的CvSQS蛋白克隆至载体pET30a,随后转化大肠杆菌TOP10,发现CvSQS被成功表达。随后该课题组以FPP为底物,加入NADPH和Mg2+进行体外实验,证明了截短的CvSQS具有将FPP转化为角鲨烯的活性。SQS缺失的苹果树叶片中β-谷甾醇和菜油甾醇的含量显著减少,实验组的菜油甾醇含量仅为对照组的37%[27],这表明了SQS在植物甾醇生产中的重要性,菜油甾醇的含量提升是提高下游油菜素内酯产量的关键。
CAS是植物甾醇合成途径中的关键调控酶之一,催化2,3-环氧角鲨烯到环阿屯醇的生物合成,属于氧化角鲨烯还原酶家族。烟草NtCAS1蛋白与拟南芥AtCAS1的序列相似性为78%[28],在缺乏内源性羊毛甾醇合酶的酵母突变体中鉴定了NtCAS1催化环阿屯醇生成的功能。一些植物如白桦Betula platyphylla Suk.和远志Polygala tenuifolia Willd中含有2个CAS基因,而重楼Paris polyphylla Smith中只含有1个CAS基因[29]。PpCAS蛋白具有761个氨基酸,具有1个DCTAE基序和4个QW(QXXXGXW和QXXXGXXXW)基序。将PpCAS基因导入羊毛甾醇合酶缺失的酵母后,重组菌株中产生了环阿屯醇,表明PpCAS可以以2,3-环氧角鲨烯为底物生成环阿屯醇,这是首次对PpCAS进行功能表征。
1.2 环阿屯醇到4-甲基-24-亚甲基胆甾-7-烯醇的生物合成途径
环阿屯醇到4-甲基-24-亚甲基胆甾-7-烯醇的合成需要经过6步反应,首先在甾醇-C24-甲基转移酶1(C24-methyltransferase 1,SMT1)的作用下生成24-亚甲基环阿屯醇,经过甾醇-4α甲基氧化酶1(sterol-4α-methyl oxidase 1,SMO1)的氧化脱去C4β位甲基得到环桉树醇,环桉树醇环异构酶(cyclopropyl sterol isomerase,CPI)接下来催化环桉树醇环丙烷环的裂解产生钝叶醇。
甾醇C14-去甲基化酶(sterol C14-demethylase,CYP51)催化钝叶醇的14α-甲基的连续氧化,依次得到14α-羟甲基、14α-醛基,最后14α-醛基以甲酸形式释放,同时生成Δ14,15双键,得到4α-甲基-5α,8,14,24(28)麦角甾三烯-3β-醇。最后在甾醇C14-还原酶(sterol C14 reductase ,C14-R)和甾醇C-8,7异构酶(sterol Δ8-Δ7isomerase,8,7-SI)的催化作用下发生C14-C15位双键还原和双键异构化分别生成4α-甲基粪甾醇和4-甲基-24-亚甲基胆甾-7-烯醇(图3-A)。
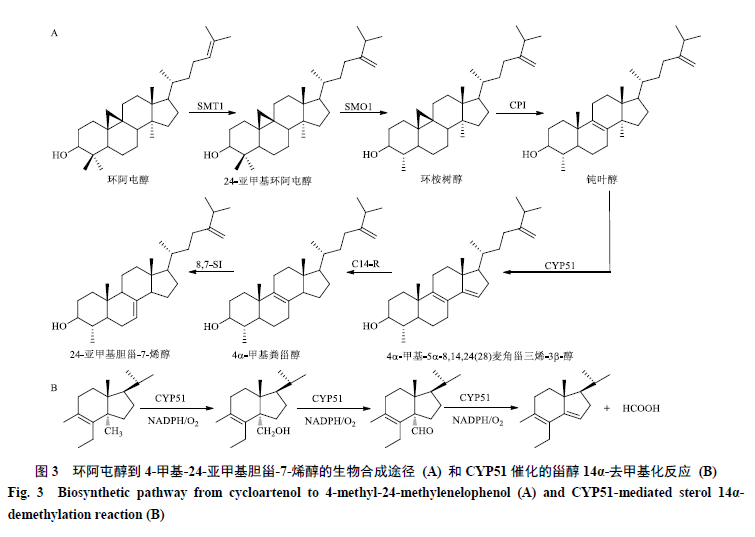
植物甾醇侧链上C24位的烷基化是由SMT催化的[30]。SMT家族的SMT1和SMT2先后在C24位引入烷基分别得到C24-甲基甾醇和C24-乙基甾醇,在有些植物如拟南芥中也存在SMT3,与SMT2发挥相同作用,都催化甾醇侧链C24位的乙基化[31-33]。1996年,植物SMT1首次从大豆中被分离出来,它在幼叶、根和茎中的表达要高于成熟叶;表达大豆SMT1的大肠杆菌可以催化羊毛甾醇C24位的甲基化[34]。重楼中有2个SMT1(PpSMT1-1和PpSMT1-2),它们都包含3段高度保守的序列:VLDVGCGIG、NSFDAVYA和VLKPGQYFAA[35]。
在大肠杆菌BL21中异源表达PpSMT1-1和PpSMT1-2,将纯化的蛋白用于体外酶促反应,PpSMT1-1可催化环阿屯醇转化为24-亚甲基环阿屯醇,而PpSMT1-2没有该催化活性。在烟草中过表达睡茄的SMT1基因,重组植株中环阿屯醇、菜油甾醇、β-谷甾醇和豆甾醇的含量大幅提高,最大增幅分别为93%、161%、587%、68%。利用可诱导植物发生多种防御反应的flg22处理拟南芥植株,与野生型相比,突变体smt2和smt3产生的活性氧显著增加,根的伸长等晚期防御反应受到抑制[36],表明SMT2和SMT3在植物免疫反应中发挥了重要作用。
在植物甾醇的生物合成中,SMO负责去除C4位上的2个甲基。SMO1去除4,4-二甲基甾醇24-亚甲基环阿屯醇C4β位的单个甲基,生成环桉树醇,SMO2则在植物甾醇生物合成的更下游步骤去除C4α位的甲基[37-39]。拟南芥中的SMO1基因(3个亚型:AtSMO1-1、AtSMO1-2和AtSMO1-3)和SMO2基因(2个亚型:AtSMO2-1和AtSMO2-2)共5个cDNA,这些AtSMO的序列中都存在3个富含组氨酸的保守序列,且表现出甾醇去饱和酶家族特有的基序[37]。拟南芥AtSMO1和酵母中的SMO(ERG25)的N端序列同源性较低,AtSMO1的N端结构域可能与识别9β,19-环丙基甾醇的特殊结构有关。异源表达AtSMO2可以恢复敲除ERG25基因的酿酒酵母菌株的生长和麦角固醇的生物合成,而异源表达AtSMO1则不可恢复,这说明了AtSMO1具有严格的底物特异性。
CPI是植物特有的酶,催化环桉树醇的环丙烷环打开,生成钝叶醇,这一反应是植物甾醇合成途径中的限速步骤之一[40]。藻类、小立碗藓、水稻、玉米、小米、杨树和拟南芥等植物中的CPI1基因均为单拷贝,它们编码的蛋白质序列相似性为48%~90%,且都具有7个跨膜结构域和6个亲水环[41],基于蛋白质同源性以及相似的结构域,推测陆生植物中CPI1蛋白的功能高度保守。拟南芥的CPI1在大肠杆菌中异源表达后,突变菌株的提取物可以将环桉树醇异构化为钝叶醇[42]。
羟胺可诱导拟南芥CPI发生点突变,通过体外测定突变体酶活,Gly28、Glu29、Gly108和Asp260对CPI活性至关重要[43],对于酶的稳定性是必需的。Men等[40]筛选到一株拟南芥CPI1的转座子插入突变体(cpi1-1),反转录聚合酶链式反应(reverse transcription- polymerase chain reaction,RT-PCR)结果表明,该突变体中没有CPI1的转录,随后对其甾醇成分进行分析,发现突变体中几种主要植物甾醇的含量几乎为零,并且积累了大量的环桉树醇,这证实了CPI基因编码环桉树醇环异构酶催化环桉树醇的环丙烷环开环的反应。
CYP51属于细胞色素P450超家族,催化甾醇14α-甲基的羟基化反应[30]。大多数CYP51有6个保守的结构域,分别为5个底物识别位点和1个血红素辅基结合位点[44]。CYP51催化的反应分3步进行,底物的14α-甲基先后被氧化为醇和醛,然后以甲酸的形式去除同时生成C14-C15位双键[45](图-3B),每一步反应都需要1个O2分子和1个NADPH[46]。小麦CYP51基因编码的蛋白与哺乳动物和真菌来源的CYP51蛋白有32%~39%的同源性,表达小麦CYP51基因的酿酒酵母可催化钝叶醇的甲基化,且比酵母内源的羊毛甾醇甲基化酶的催化效率更高[47]。拟南芥中有2个CYP51基因(CYP51A1和CYP51A2),序列分析和分子互补实验证明CYP51A1是1个假基因,cyp51A2突变体中积累了大量的钝叶醇,菜油甾醇和β-谷甾醇含量减少[48-49]。
1.3 4-甲基-24-亚甲基胆甾-7-烯醇到菜油甾醇的生物合成途径
4-甲基-24-亚甲基胆甾-7-烯醇到菜油甾醇的合成需要经过4步反应。首先,SMO2去除4-甲基-24-亚甲基胆甾-7-烯醇C4位的第二个甲基生成表甾醇,而后经甾醇C5-去饱和酶(sterol C5-desaturase,SC5D)、甾醇Δ7-还原酶(sterol Δ7-reductase,7-DR)和甾醇侧链还原酶1(sterol side chain reductase 1,SSR1SC5D1)的脱氢和还原作用依次生成5-脱氢表甾醇、24-亚甲基胆固醇和菜油甾醇(图4)。
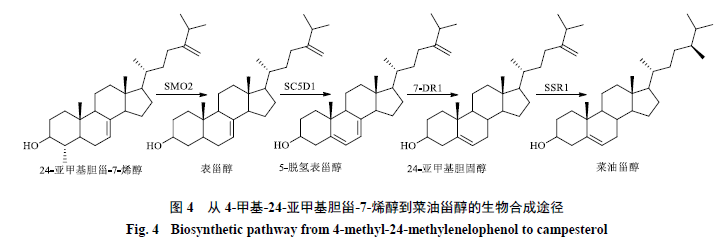
Gachotte等[50-51]利用甲基磺酸乙酯对拟南芥进行诱变处理,分离出突变株ste1-1,其Δ7-甾醇含量增加,而Δ5-甾醇含量减少,表明该突变株缺乏SC5D活性。用酿酒酵母中编码SC5D蛋白的ERG3基因转化ste1-1突变株的根,其愈伤组织中Δ5-甾醇含量增加了1.7~2.8倍,Δ7-甾醇含量减少,说明ste1-1中Δ7-甾醇的积累是由SC5D活性缺陷引起的,而后从拟南芥中分离了SC5D。拟南芥中的DWF7基因是STE1的等位基因,不同植物的SC5D在序列和特定结构域高度保守,具有紧密排列的组氨酸残基HX3H[52]。黄色荧光蛋白融合实验表明,拟南芥的SC5D主要定位于内质网,在质膜和脂质颗粒中也有表达[53]。番茄中存在2种SC5D蛋白(SC5D1和SC5D2),二者氨基酸序列相似性为88%[2]。SC5D1参与菜油甾醇、β-谷甾醇、和豆甾醇的生物合成,而SC5D2则在胆固醇的生物合成途径中发挥作用。
制菌霉素类多烯化合物的抗真菌作用依赖于麦角固醇中的C7位不饱和键,酵母中的ERG2催化麦角固醇中C7位双键的生成,因此ERG2基因敲除的酿酒酵母突变株表现出对制菌霉素的高度耐药性。通过拟南芥cDNA文库在酿酒酵母中的异源表达筛选出了拟南芥中的7-DR1,从105个转化子中筛选出1个单克隆,其对制菌霉素的抗性达到80 μg/mL,说明该cDNA编码的酶可催化麦角固醇C7位双键的还原[54],将拟南芥的7-DR1基因导入酿酒酵母菌株,首次实现了菜油甾醇在酿酒酵母中的生物合成。将拟南芥的7-DR1基因与黄色荧光蛋白融合并转化酿酒酵母W303-B1菌株,显示7-DR1蛋白主要定位于内质网,并且观察到Δ5,7-甾醇如麦角固醇的C7位双键被还原[53]。
SSR1催化菜油甾醇生物合成过程中的最后一步的C24位亚甲基还原反应。SSR1蛋白由DWF1基因编码,T-DNA插入突变的拟南芥dwf1突变株中24-亚甲基胆固醇的含量比野生型提高了12倍,而几乎没有菜油甾醇的产生[55]。SSR1蛋白中N段的27~43号残基含有跨膜结构域,可使其定位至内膜系统。在跨膜结构域下游是FAD结合结构域,FAD是SSR1的C24位亚甲基还原活性中的关键辅酶。缺失DWF1基因的植株中菜油甾醇和其下游的油菜素内酯的含量都显著下降,产生植株短小、叶子呈现深绿色、繁殖能力下降等一系列表型。
1.4 4-甲基-24-亚甲基胆甾-7-烯醇到β-谷甾醇和豆甾醇的生物合成途径
4-甲基-24-亚甲基胆甾-7-烯醇生成β-谷甾醇需要经过5步反应。SMT2在4-甲基-24-亚甲基胆甾-7-烯醇的C24位引入第2个甲基,生成4-甲基-24-亚乙基-7-胆甾烯醇,将代谢流引入C24-乙基甾醇合成途径。随后4-甲基-24-亚乙基-7-胆甾烯醇经过SMO2的转化脱去C4位甲基生成Δ7-燕麦甾醇,SC5D1催化Δ7-燕麦甾醇的C5-C6位脱氢生成5-脱氢燕麦甾醇,后者在7-DR1和SSR1的作用下依次发生C7-C8位和C24-C28位双键还原,生成β-谷甾醇。最终β-谷甾醇的C22位和C23位在甾醇C22-去饱和酶(sterol C22-desaturase,SC22D)的作用下脱氢生成豆甾醇(图5)。
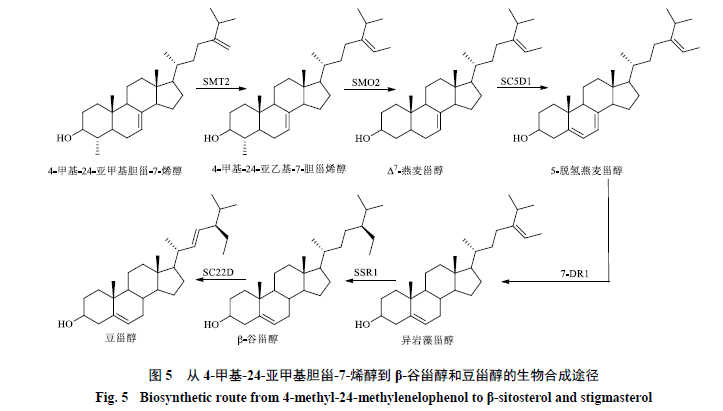
豆甾醇与β-谷甾醇的结构相似,区别在于豆甾醇侧链上C22位和C23位之间存在双键,SC22D催化β-谷甾醇向豆甾醇转化。拟南芥中有4个编码SC22蛋白的基因(CYP710A1~A4),在不同组织和器官中表达水平有较大差异。CYP710A1在根、叶和花等各种器官内的表达水平较高,在茎和成熟长角果中的表达水平非常低,CYP710A2在长角果中的表达水平也较低,CYP710A3在茎中特异性表达,CYP710A4在根中的表达较弱。在昆虫细胞中利用杆状病毒表达载体系统分别表达拟南芥的CYP710A1和CYP710A2以及番茄的CYP710A11,提取昆虫的微粒体部分进行体外催化实验,发现这些重组蛋白均可以将β-谷甾醇底物转化为豆甾醇[56],CYP710A2和CYP710A11对β-谷甾醇的亲和力更高。
此外,同样在昆虫细胞中利用杆状病毒表达载体系统表达编码苔藓SC22D的CYP710A3和CYP710A4,验证了其编码蛋白的C22位去饱和活性[57]。在本氏烟草中将番茄SC22D与绿色荧光蛋白融合表达,发现该融合蛋白定位于内质网[58]。进一步的研究表明,番茄SC22D中的N端跨膜螺旋(transmembrane-helix domain,TMH)结构域可以使其靶向内质网。将完整的番茄SC22D和缺失TMH结构域的SC22D在本氏烟草中分别表达,发现表达完整SC22D的烟草中豆甾醇含量增加,而缺失TMH结构域的SC22D没有去饱和酶活性。不同植物的SC22D中TMH结构域的长度和序列保守性均有较大差异,但都含有大量的苏氨酸和丝氨酸残基[58]。
2 植物甾醇衍生物的生物合成途径
2.1 甾醇酯、甾醇糖苷以及酰基甾醇糖苷的生物合成途径
甾醇酯是由游离甾醇的C3位羟基被酯化而形成的,这一过程由甾醇酰基转移酶(SATs)催化(图6)。通常根据酰基供体的不同将SATs分为磷脂甾醇酰基转移酶(PSATs)和酰基辅酶A甾醇酰基转移酶(ASATs)2类[59]。磷脂和中性脂质,包括二酰基甘油和三酰基甘油,都可以作为酰基供体参与甾醇酯的生物合成[60-62]。来源于拟南芥的甾醇酰基转移酶AtPSAT催化酰基从磷脂转移到各种游离甾醇的C3位[63],微粒体实验表明AtPSAT对不同游离植物甾醇具有底物特异性,其底物偏好性依次为胆固醇、菜油甾醇、β-谷甾醇和豆甾醇。
从酰基供体来考察,AtPSAT优先利用磷脂酰乙醇胺,其次是磷脂酰胆碱,几乎不能利用磷脂酸。在烟草和拟南芥中过表达AtPSAT1基因,检测到其mRNA的大量积累,但是在叶子和种子中并没有检测到总甾醇和甾醇酯水平的升高,这可能与甾醇稳态有关[64]。水稻OsPSAT基因的表达水平与脱落酸含量和HMGR基因的表达呈正相关,当水稻处于干旱胁迫条件下时OsPSAT基因的表达水平升高,提示甾醇酯可能与植物的应激反应相关[65]。番茄的PSAT(SlPSAT1)与AtPSAT的序列相似性为75%,在缺乏AtPSAT基因的拟南芥突变株中表达SlPSAT1基因可逆转突变株中观察到的早期衰老表型,并使甾醇酯产量恢复至野生型水平[59]。
拟南芥的AtASAT定位于内质网,而番茄的SlASAT1则位于质膜,二者都对环阿屯醇有明显的底物偏好性[59]。SlPSAT1基因和SlASAT1基因响应不同外界刺激(脱落酸、茉莉酸甲酯、水杨酸、渗透、盐和冷胁迫等)时的反应有所不同,SlPSAT1在胁迫条件下表现出明显的表达水平差异,而SlASAT1在不同处理下表达水平几乎保持不变[59]。甾醇酯可以在甾醇酯酯酶的作用下转化为游离甾醇,以维持植物的甾醇稳态。
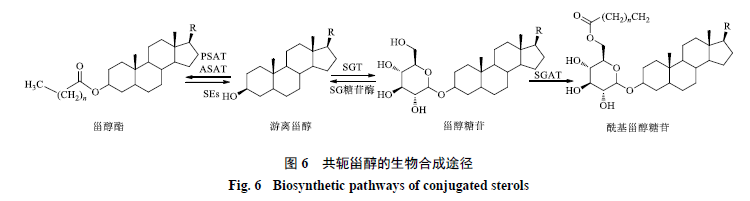
甾醇糖苷由甾醇糖基转移酶(sterol glycosyltransferase,SGT)催化游离甾醇的C3位羟基糖基化而产生[66-67](图6)。SGT是糖基转移酶家族的成员,其C端含有1个糖结合域(AIIANPPAY)和1个甾醇结合域(VVHHGGAG)[68-70]。目前,多种植物的SGT已经被克隆并在大肠杆菌中进行了异源表达,多数SGT都优先利用尿苷二磷酸葡萄糖作为糖基供体。研究表明,SGT的活性受到细胞膜脂质环境的影响,带负电荷的磷脂可以增强SGT的活性,但这一现象的机制目前还不清楚[71-74]。将拟南芥和燕麦中的膜结合SGT分别在大肠杆菌中表达并纯化,体外实验证实这2种酶可以催化相关膜甾醇的糖基化[75]。
睡茄叶子存在一种胞质SGT,其仅以β-谷甾醇、豆甾醇和薯蓣皂苷元等C3羟基甾醇作为底物,且其催化5-烯甾醇的糖基化效率比5α-H甾醇更高[76]。拟南芥中的2个SGT功能有所不同,其中At3g07020基因编码的UGT80A2负责种子中主要甾醇糖苷的生产,而At1g43620基因编码的UGT80B1在甾醇糖苷的合成过程中起辅助作用[77-78]。棉花中的GhSGT1主要利用胞质甾醇,而GhSGT2主要以质膜甾醇为底物[79]。非离子表面活性剂Triton X-100对GhSGT1的活性有抑制作用,但可显著增强GhSGT2的活性。添加NaCl后,GhSGT1的活性要强于GhSGT2,而在添加MgCl2后则相反。真核生物的甾醇稳态依赖于游离甾醇和共轭甾醇的相互转化,甾醇糖苷可以在糖苷酶的作用下发生糖苷水解而生产游离甾醇。
甾醇糖苷中糖基的C6位可以被脂肪酸酯化形成酰基甾醇糖苷,这一过程由甾醇糖苷酰基转移酶(steryl glucoside acyltransferase,SGAT)催化(图6)。Forsee等[80]分离并纯化了棉花中的SGAT,将该酶与14C标记的磷脂、3H标记的甾醇糖苷一同在体外做催化实验,在产物中检测到了14C, 3H标记的酰基甾醇糖苷,证明了磷脂可以作为甾醇糖苷向酰基甾醇糖苷转化的酰基供体。来自茄叶的膜结合SGAT具有明显的底物特异性,在所有测试的甾醇糖苷底物中,当以胆固醇葡萄糖苷为底物时该酶的活性最高,以其他甾醇糖苷如β-谷甾醇葡萄糖苷、豆甾醇葡萄糖苷和胆固醇半乳糖苷等为底物时酰基化效率较低[81]。进一步的研究发现,酰基来源可以是不同类型的磷酸甘油酯,如磷脂酰乙醇胺、磷脂酰胆碱、磷脂酰肌醇和磷脂酰甘油等。
2.2 油菜素内酯的生物合成途径
目前已在不同植物中鉴定出几十种不同的油菜素内酯类植物激素,芸苔素内酯(brassinolide,BL)是第1个被鉴定的油菜素内酯类化合物,也是研究这类植物激素的生物合成途径的模式化合物。以BL为例介绍油菜素内酯类化合物的生物合成途径。BL的合成途径呈复杂的网络状结构,根据C22位发生羟基化的时间不同,可以将从菜油甾醇到BL的上游途径中分为早期羟基化途径和晚期羟基化途径。根据C6位的氧化时间,又可以将菜油甾醇到BL的下游途径分为早期C6氧化途径和晚期C6氧化途径(图7)。
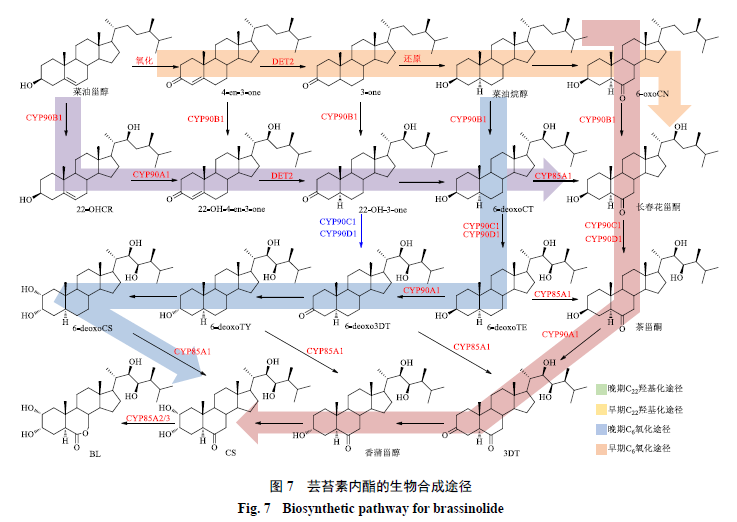
早期羟基化途径的第1步是菜油甾醇的C22位发生羟基化,生成(22S)-22-羟基菜油甾醇[(22S)-22-hydroxycampesterol,22-OHCR]。22-OHCR的C3位羟基氧化,C4、C5位双键异构化后转化为(22S,24R)
-22-hydroxyergost-4-en-3-one(22-OH-4-en-3-one),后者进一步经过C4、C5位双键加氢还原和C3位羰基加氢还原依次生成(22S,24R)-22-hydroxy-5α-ergostan-3-one(22-OH-3-one)和6-脱氧长春花甾酮(6-deoxo-cathasterone,6-deoxoCT)[82]。
在晚期C22羟基化途径中,菜油甾醇依次经过C3位羟基脱氢,C4、C5位双键异构化,C4位双键加氢还原和C3位羰基还原依次转化为(24R)-ergost-4-en-3-one(4-en-3-one)、(24R)-5a-ergostan-3-one(3-one)和菜油烷醇[83]。菜油烷醇的C6位氧化为羰基,生成6-氧菜油烷醇(6-oxocampestanol,6-oxoCN)。6-oxoCN的C22位发生羟基化是晚期C22羟基化途径的最后一步,生成长春花甾酮。
晚期C6氧化途径与早期C6氧化途径的合成顺序高度相似,都以菜油烷醇作为途径的起始物,区别在于C6位氧化反应发生的时间不同(图7)。在早期C6氧化途径中,菜油烷醇的C6位首先被氧化,生成6-oxoCN;然后C22和C23位依次发生羟基化,转化为长春花甾酮和茶甾酮[84]。茶甾酮经C3位脱氢和还原、C2位羟基化依次生成3-脱氢茶甾酮(3DT)、香蒲甾醇和油菜素甾酮(CS)。CS的B环发生Baeyer-Villiger氧化重排,最终被转化为BL[85-86]。
而在晚期C6氧化途径中,菜油烷醇经过羟基化、脱氢、还原等反应转化为早期C6氧化途径中相应中间体的C6脱氧形式,即6-deoxoCT、6-脱氧茶甾酮(6-deoxoTE)、6-脱氧-3-脱氢茶甾酮(6-deoxo3DT)、6-脱氧香蒲甾醇(6-deoxoTY)和6-脱氧油菜素甾酮(6-deoxoCS),C6位的氧化发生在最后一步生成CS。相较于C6早期氧化途径,植物中晚期C6氧化途径中的中间产物浓度更高,因此被认为是主要的BL合成途径。BL生物合成途径中的酶并不完全清晰,目前已解析的酶主要包括CYP90B1(DWF4)、CYP90A1、CYP90C1、CYP90D1、CYP85A1、CYP85A2和CYP85A3[87-92]。
DWF4基因编码的类固醇22α-羟化酶CYP90B1[93],负责催化BL生物合成过程中如菜油甾醇、4-en-3-one、3-one、菜油烷醇、6-oxoCN等中间体的C22位羟基化[86],也是菜油甾醇合成BL途径中的第1个限速酶[88]。T-DNA插入的dwf4突变植株表现出典型的油菜素内酯缺陷表型,这种表型可以被BL和C22羟基化的油菜素内酯中间体恢复,但是不能被其他植物激素恢复[94-96]。
DWF4蛋白定位于内质网,RT-PCR和GUS染色显示DWF4主要在根尖、茎尖、花、幼苗、腋芽和角果中表达[97]。在拟南芥中过表达DWF4后促进了植株的生长,同时BL产量增加。研究表明,拟南芥同源域亮氨酸拉链蛋白1可识别并结合到DWF4的启动子区域,同时联合油菜素内酯合成途径下游的转录因子BES1共同抑制DWF4的表达[98]。染色体免疫共沉淀和凝胶迁移滞后实验发现转录因子TCPI可直接结合到DWF4启动子区的2个GGNCC靶序列上,提高CYP90B1的表达水平[99]。
CYP90C1和CYP90D1都属于C23羟基化酶,可负责特异性催化BL合成途径中C22羟基化中间体的C23位羟基化。体外催化实验表明,含有CYP90C1或CYP90D1的微粒体和纯化的拟南芥CYP450还原酶可将6-deoxoCT和长春花甾酮分别转化为6-deoxoTE和茶甾酮[90]。此外,除了C22羟基化途径和C6氧化途径外,还存在另外一条绕过菜油烷醇和6-deoxoTE的新途径,即22-OH-3-one的C23位在CYP90C1或CYP90D1的作用下发生羟基化,直接转化为6-deoxo3DT。
CYP85A1可催化晚期C-6氧化途径中不同中间体,如6-deoxoCS、6-deoxoTY、6-deoxo3DT和6-deoxoTE上C6位的氧化反应,表明这一氧化酶具有良好的底物杂泛性。番茄和拟南芥中6-deoxoCS向CS的转化是由细胞色素P450的CYP85A家族催化的[100-101]。目前,已经在水稻、葡萄、豌豆、番茄和拟南芥等多种植物中分离了CYP85A1。将豌豆的CYP85A1和CYP85A6分别在酿酒酵母中进行异源表达[102],重组酿酒酵母与6-deoxoCS共同培养时,CYP85A1和CYP85A6均可将6-deoxoCS转化为CS。此外,将表达CYP85A1的酿酒酵母与CS一起培养时,在代谢物中还产生了微量的BL,说明CYP85A1也具有一定的催化Baeyer-Villiger反应产生BL的活性。而表达CYP85A6的重组酿酒酵母则不能将CS转化为BL。
从CS到BL的转化过程中,在CS的C6和C7位之间发生Baeyer-Villiger氧化,插入1个氧原子产生7-氧代-内酯B环,这一反应也是BL生物合成的限速步骤之一。异源表达拟南芥CYP85A2基因的酿酒酵母可以催化CS转化为BL[101,103-104]。将CS和氘标记的CS添加到导入拟南芥CYP85A2的酵母培养基中,在代谢物中可同时检测到未标记和标记的BL[104],证实了CYP85A2具有催化Baeyer-Villiger氧化反应的活性。在酵母中进行异源表达番茄中的CYP85A3基因,发现重组酵母菌株将大部分6-deoxoCS转化为CS,同时也观察到BL的产生[92],说明CYP85A3也参与BL的生物合成。
3 合成生物学在植物甾醇及其衍生物生产中的应用
利用合成生物学技术高产植物甾醇的前提是对其生物合成途径进行挖掘。近年来,随着对植物甾醇生物合成途径研究的不断深入以及现代合成生物学技术的快速发展,已经成功在微生物细胞中成功构建了植物甾醇的合成途径。Du等[105]在敲除内源C22去饱和脱氢酶编码基因的解脂耶氏酵母中导入不同来源(水稻、褐家鼠、非洲爪蟾)的7-脱氢胆固醇还原酶编码基因DHCR7,实现了菜油甾醇在酵母中的发酵生产。
在这些基因中,表达非洲爪蟾来源DHCR7基因的菌株中菜油甾醇产量最高,摇瓶产量达到106 mg/L。随后通过高密度补料分批发酵,以葵花籽油为碳源,最终菜油甾醇最高产量达到(453.0±24.7)mg/L。将来自不同物种的DHCR7基因在解脂耶氏酵母中表达以提高重组酵母中菜油甾醇的产量,发现来源于斑马鱼的DHCR7基因是合成菜油甾醇的最佳候选酶[106]。
过表达斑马鱼DHCR7基因、柠檬酸裂解酶和过氧化物酶体酰基辅酶A氧化酶2的重组酵母,在5 L发酵罐中的菜油甾醇最高产量达到942 mg/L。在高产角鲨烯的酵母底盘细胞中表达外源基因DHCR7,通过基因来源筛选、启动子以及发酵条件优化,最终菜油甾醇的产量达到了(916.88±11.23)mg/L[107]。由此可见,通过合成生物学技术,将不同来源的基因在微生物细胞中有效整合,构建植物甾醇的人工生物合成途径,从而实现高值植物甾醇的发酵生产是可行的。
4 结语与展望
植物甾醇及其衍生物具有降血糖、调血脂[108]、降胆固醇、抗肿瘤、抗炎、抗氧化和预防心血管疾病等生理活性,其生物合成途径在近年来也得到了较为深入的研究,本文对这些化合物的生物合成途径进行了总结。在植物甾醇的完整生物合成途径及调控机制方面还存在一些尚待解决的问题。
首先,在BL的生物合成途径中,有一些反应的酶还没有被发现,如菜油甾醇到4-en-one、3-one到6-oxoCN、22-OH-3-one到6-deoxoCT、6-deoxoCT到6-deoxoCS以及3DT到CS的生物合成途径中的酶,需要进一步对这些未知的生物合成途径进行探究,通过筛选BL合成突变体,分离并鉴定新的酶,分析其功能和催化的反应步骤。
第二,在植物甾醇的生物合成途径中,有的酶可以催化不同底物发生相同的反应,如CYP85A1可催化BL生物合成途径中多个中间体的C6位氧化反应;CYP90B1催化晚期C22羟基化途径中多个底物的C22位羟基化;另外,不同的酶可以催化相同的反应过程,如SMT2和SMT3都可催化侧链C4位的乙基化。这些酶的功能比较复杂,需要进一步去探究它们的调控机制。
第三,一些植物甾醇的生物合成途径比较长,且具有高度的复杂性,这种情况下酶的反应往往不完全,易产生限速步骤,有待于进一步优化生物合成途径,以提高产量。
通过合成生物学技术将植物甾醇的生物合成途径导入异源底盘微生物中,只需要利用廉价易得的碳源就可实现植物甾醇的从头合成,不仅能够高效获得目标产物,而且环境友好,具有广阔的市场发展前景及社会经济效益。对植物甾醇生物合成途径进行系统的研究将会为更好地利用现代合成生物学技术获得目标甾醇提供优选方案。随着对更多植物甾醇生物合成途径的解析和更多合成酶的发现与表征,以及关键催化酶的活性提升和底盘细胞的代谢工程改造,有望进一步提升植物甾醇在微生物细胞中的合成效率和产量,使植物甾醇的规模化发酵生产成为可能。
版权声明:本文内容由互联网用户自发贡献,该文观点仅代表作者本人。本站仅提供信息存储空间服务,不拥有所有权,不承担相关法律责任。如发现本站有涉嫌抄袭侵权/违法违规的内容, 请发送邮件至 2303805254@qq.com,本站将立刻删除。